
Международная группа палеонтологов выпустила свежий обзор данных, касающихся появления в палеонтологической летописи первых представителей самого крупного типа животных — членистоногих. Сразу несколько дисциплин, включая молекулярную филогенетику и тафономию (науку о закономерностях захоронения ископаемых остатков), приводят к выводу, что реальным временем возникновения членистоногих был рубеж эдиакарского и кембрийского периодов, когда произошел так называемый «кембрийский взрыв». Авторы обзора отвергают конкурирующую гипотезу «долгой скрытой докембрийской эволюции», согласно которой главные ветви животных, в том числе и членистоногих, возникли гораздо раньше.
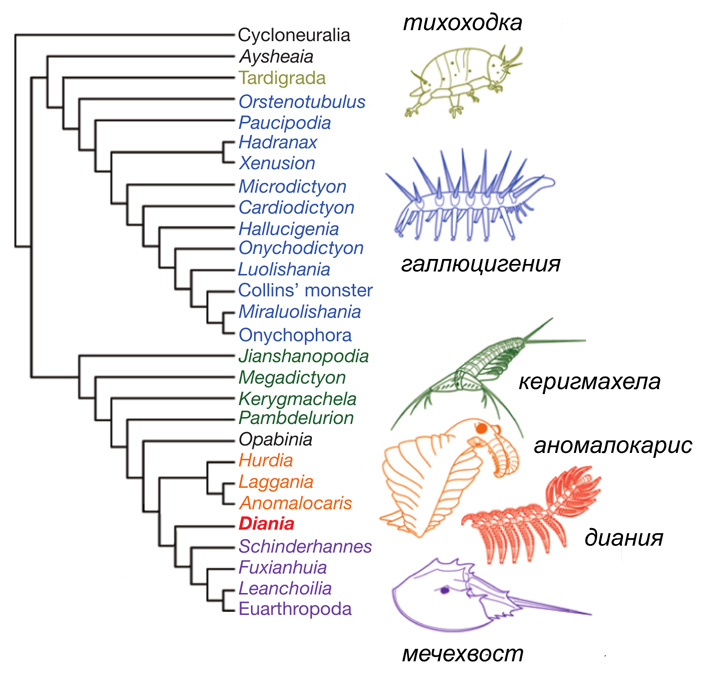
Рис. 1. Эволюционное древо панартропод, сопровожденное изображениями некоторых членов этой группы. Подробные объяснения — в тексте. Детальная структура приведенного древа в данном случае неважна, это просто одна из версий. Среди изображенных животных — два современных (тихоходка и мечехвост) и три ископаемых, из которых галлюцигения (Hallucigenia) близка к онихофорам, а керигмахела (Kerygmachela), аномалокарис (Anomalocaris) и диания (Diania) относятся к обсуждаемым ниже «стволовым эуартроподам». Стоит упомянуть, что керигмахела получила видовое название в честь великого датского философа Сёрена Кьеркегора — Kerygmachela kierkegaardi. Иллюстрация из статьи D. A Legg et al., 2013. Lobopodian phylogeny reanalised, с изменениями
Хозяева Земли
Говорят, что знаменитый английский биолог Джон Холдейн (John Burdon Sanderson Haldane) однажды оказался в компании богословов, и они задали ему вопрос: какие черты Творца обнаруживаются при изучении природы творения? Холдейн, известный своими материалистическими взглядами, проворчал: «Неумеренная увлеченность жуками» («An inordinate fondness for beetles»; цитируется по статье: G. E. Hutchinson, 1959. Homage to Santa Rosalia or why are there so many kinds of animals?). Этот шуточный ответ был, однако, основан на биологических фактах. Широко известно, что жуки, или жесткокрылые, — это самый многочисленный отряд во всем животном царстве. Если, например, сравнить жуков и млекопитающих, окажется, что число известных на данный момент видов жуков превосходит число видов млекопитающих в 72 раза. И это несмотря на то, что млекопитающие — не отряд, а класс, т. е. группа значительно более высокого уровня.
Сказанное о жуках можно распространить и на их эволюционных родственников — от близких до дальних. Жуки относятся к насекомым, которые, в свою очередь, входят в тип членистоногих. Разнообразие этого типа поистине грандиозно. По современным подсчетам, больше 80% всех видов многоклеточных животных — это членистоногие. Другой настолько успешной эволюционной ветви просто нет в природе. Замечательная книга Эдварда Уилсона (Edward Osborne Wilson), значительная часть которой рассказывает о возникновении социальных систем у членистоногих, недаром называется «Хозяева Земли».
Неудивительно, что эволюция членистоногих занимает многих исследователей. Дело облегчается тем, что палеонтологическая летопись членистоногих сравнительно богата. Более того, ее «чтение» — работа достаточно благодарная. Согласно принципу актуализма, ключом к пониманию прошлого служит настоящее. В частности, любые выводы, касающиеся ископаемых животных, должны быть в конечном счете обязательно основаны на данных о животных современных — иначе исследователю будет просто не на что опереться. В случае с членистоногими этот принцип работает прекрасно, потому что в распоряжение ученых попадает огромное количество как современных, так и ископаемых форм, очень разнообразных, но в то же время устроенных более-менее по одному плану. Это — идеальная ситуация для палеонтолога, и естественно, что исследователи ископаемых членистоногих пользуются ей сполна (см., например: Палеоэнтомология в России).
Членистоногие — группа организмов, очень подходящая для проверки всевозможных гипотез, касающихся хода эволюции. Например, действительно ли большинство современных крупных групп животных возникло во время кембрийского взрыва, или их внезапное появление в начале кембрийского периода связано с образованием твердых скелетов, а истинные эволюционные «корни» лежат гораздо глубже? Попытки выяснить это, основываясь на материале по членистоногим, уже предпринимались (см. Членистоногие подтверждают реальность кембрийского взрыва, «Элементы», 17.11.2013). Но эволюция членистоногих, особенно ранняя, крайне интересна и сама по себе — как история, героями которой являются многие совершенно необычные с нашей современной точки зрения живые существа. Палеонтология XXI века успела сделать много открытий в этой области. Впрочем, и нерешенных вопросов тут, как всегда, еще хватает.
Усики, жвалы и хелицеры
По современным данным, тип членистоногих состоит из двух крупных эволюционных ветвей.
Одна ветвь — хелицеровые (Chelicerata), куда относятся паукообразные, мечехвосты и морские пауки. У них нет усиков, зато есть хелицеры — околоротовые придатки, оканчивающиеся клешнями, крючками или стилетами. Большинство хелицеровых, во всяком случае крупных, — хищники, потому что для других способов питания их ротовой аппарат подходит мало.
Вторая ветвь членистоногих называется жвалоносными (Mandibulata). В их ротовой аппарат входят челюсти — в том числе мандибулы, которые иначе называются жвалами, — представляющие собой не что иное, как сильно видоизмененные ходильные ноги. Этим жвалы принципиально отличаются от хелицер, которые не имеют с ногами ничего общего. Ротовой аппарат жвалоносных, оснащенный набором челюстей, позволяет осваивать предельно разнообразные способы питания, что мы и видим на примере членов этой группы — многоножек, ракообразных и особенно насекомых. Кроме того, для жвалоносных очень характерны усики, или антенны — одна или две пары. Как уже упоминалось, у хелицеровых усиков нет.
Строго говоря, тип, состоящий из жвалоносных и хелицеровых, называется Euarthropoda (эуартроподы, «настоящие членистоногие»). Это название, более точное, чем просто «членистоногие», предложил в 1904 году знаменитый английский зоолог Рэй Ланкестер (Sir Edwin Ray Lankester). Среди признаков эуартропод — четкое деление тела на отделы (тагмы) и присутствие членистых конечностей, внутри которых есть суставы.
Животные, у которых членистых конечностей нет и никогда не было, не относятся к эуартроподам, даже если по многим другим признакам они к ним близки. Существует две группы таких животных: онихофоры — наземные существа, которых иногда называют «бархатными червями» (velvet worms), — и тихоходки, мельчайшие водяные жители (см., например: Укорочение тела тихоходок связано с потерей Hox-генов, «Элементы», 04.03.2016). И у онихофор, и у тихоходок тело сегментировано почти так же, как у членистоногих, но конечности нечленистые. Поэтому диагнозу типа Euarthropoda, равно как и буквальному значению слова «членистоногие», они не соответствуют.
Тем не менее онихофоры и тихоходки, безусловно, являются близкими родственниками эуартропод. В конце XX века эуартропод, онихофор и тихоходок формально объединили в надтип Panarthropoda (букв. «все членистоногие»). Панартроподы — огромная эволюционная ветвь, которая, в свою очередь, входит в группу линяющих животных (Ecdysozoa; см., например: Китайские палеонтологи нашли древнейшего головохоботного червя, «Элементы», 04.06.2014). Таково положение членистоногих в системе животного мира.
«Стволовые эуартроподы»
До сих пор, говоря о членистоногих и их родственниках, мы упоминали только современные группы. Однако для полного понимания эволюционных событий, сформировавших самый многочисленный тип животного царства, просто необходимо учитывать палеонтологическую летопись, и прежде всего — летопись кембрийского периода, когда сформировались почти все крупные эволюционные ветви животных. В кембрии встречаются интереснейшие живые существа, совершенно не похожие на своих современных родственников (многих из них нельзя отнести ни к какому современному типу — по крайней мере, без явных натяжек). Трудность в том, что «мягкотелые» существа, у которых нет минеральных скелетов или раковин, сохраняются в ископаемом состоянии относительно редко, и к членистоногим это, увы, тоже относится. Для их захоронения нужны лагерштетты — глинистые осадочные породы, образующиеся в малокислородных условиях (там, где кислорода много, в грунт проникают роющие животные, которые сразу нарушают структуру формирующегося осадка; см. А. Ю. Журавлев, 2014. Ранняя история Metazoa — взгляд палеонтолога). Особой известностью в качестве источника кембрийских ископаемых, заключенных в лагерштеттах, пользуются канадские сланцы Бёрджесс (Burgess Shale), где раскопки ведутся уже больше ста лет. Фауна сланцев Бёрджесс настолько знаменита, что местонахождения с подобным типом сохранности обозначаются в научных статьях аббреавиатурой BST — Burgess Shale-type, «тип сланцев Бёрджесс». Палеонтология в наши дни развивается быстро, и местонахождений BST известно много, причем в некоторых из них степень сохранности просто поразительна. Это позволило неплохо изучить древнейших родственников членистоногих и, что называется, пролить свет на их происхождение. Правда, открывшуюся картину никак не назовешь простой. Но тем она интереснее.
Бросим взгляд на эволюционное древо (рис. 2). По молекулярным данным, которые в данном случае неплохо согласуются с палеонтологическими, ближайшие «внешние» родственники эуартропод, дожившие до наших дней — это онихофоры. В палеонтологической летописи онихофоры — или, во всяком случае, близкие к ним формы — тоже присутствуют. Правда, в кембрийском периоде они были еще не наземными, а морскими (см. Aysheaia). Это гусеницеобразные существа, которые передвигались по дну на множестве коротких ножек, оканчивающихся коготками. Вполне вероятно, что общие предки эуартропод и онихофор были на них похожи.

Рис. 2. Упрощенное эволюционное древо членистоногих и их родственников. Тихоходки, положение которых до сих пор остается спорным и которые в любом случае представляют собой крайне специализированную боковую ветвь, сюда не включены. Оранжевая стрелка указывает на представителя «стволовых эуартропод». У него можно видеть плавательные выросты на туловище, фасеточные глаза и длинные, мощные членистые околоротовые придатки (об этих признаках см. ниже в тексте)
Однако онихофоры вряд ли являются непосредственными предками эуартропод (во всяком случае, эуартропод в строгом и традиционном смысле этого слова). Вот тут-то и начинается самое интересное. На реальном эволюционном древе между ветвью онихофор и ветвью эуартропод находится целая совокупность весьма оригинальных полностью вымерших ветвей, члены которых обладали отдельными «эуартроподными» признаками, но еще не набрали их полного комплекса, какой мы видим у паука, сороконожки или мухи. Эти ветви можно было бы назвать переходными, но надо помнить, что для своей среды и своего времени каждое относящееся к ним животное было вполне завершенным организмом, отлично вписанным в определенную экологическую нишу (иначе бы оно не попало в палеонтологическую летопись). Хотя, конечно, с нашей современной точки зрения многие члены этих «переходных» ветвей выглядят крайне странно.
В современной научной литературе упомянутую совокупность эволюционных ветвей принято называть «стволовой группой эуартропод», или просто «стволовыми эуартроподами» (stem-Euarthropoda). Честно говоря, такое обозначение может изрядно запутать: ведь в том-то и дело, что животные, на которых оно распространяется, к эуартроподам в узком смысле этого слова не относятся (см. J. Ortega‐Hernandez, 2014. Making sense of ‘lower’and ‘upper’ stem‐group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848). Однако — тут нам поневоле придется употребить предложение, насыщенное терминами — кладистическая систематика, господствующая в современной биологии почти безраздельно, категорически запрещает вводить парафилетические таксоны. Если говорить попросту, это означает, что группа организмов, охватывающая не единственную эволюционную ветвь, а целый эволюционный уровень, не может считаться единицей классификации и иметь собственное научное название (в частности, латинское). Поскольку в данном случае такая группа налицо и как-то называть ее все же надо, исследователи используют временное обозначение, которое было сочтено корректным по формальным соображениям. Мы тоже — куда деваться — будем этим обозначением пользоваться, оставляя его в кавычках и помня, что с точки зрения классической зоологии «стволовые эуартроподы» — это не эуартроподы.
Положение «стволовых эуартропод» на эволюционном древе показано на рис. 2. Если на время оставить в стороне строгую терминологию, можно сказать, что «стволовые эуартроподы» — это эволюционный уровень, охватывающий часть древа выше онихофор и ниже эуартропод в узком смысле. Что же на этом эволюционном уровне происходило?
А происходило там много интересного. «Стволовые эуартроподы» могут поразить чье угодно воображение — настолько необычен и многообразен их облик (см. рис. 1 и 3). Их членистое туловище часто украшали ряды жаброподобных, крылоподобных или лезвиеподобных выростов, способных служить своего рода плавниками. Появлялись хитиновые головные щиты, которые в некоторых группах становились двустворчатыми. Ходильные конечности менялись во всем диапазоне от примитивных «сосочков» (или их отсутствия) до сложно устроенных длинных ног, членистых, а иногда и двуветвистых, как у ракообразных. Невероятным разнообразием отличались околоротовые придатки: членистые или нечленистые, зачаточные или очень мощные, короткие или длинные, хватательные или фильтрующие, иногда ветвящиеся, а иногда с клешнями, крючками, шипами или щупальцами. Наконец, у ряда форм возникли огромные сложные фасеточные глаза, примерно такие же, как у современных насекомых, и иногда сидящие на стебельках. И все эти признаки вступали между собой во множество сочетаний, часто неожиданных для современных зоологов.

Рис. 3. Панартроподы из сланцев Бёрджесс. А — близкая к современным онихофорам Aysheaia. B — Opabinia, обладатель плавниковых выростов, фасеточных глаз и членистого хобота, в данном случае подогнутого под туловище. C — Hurdia, от которой сфотографирован только ротовой аппарат с концентрическим расположением хитиновых зубцов (у настоящих членистоногих ничего подобного не бывает). D — Anomalocaris. Хорошо видны длинные членистые околоротовые придатки. E — еще один Anomalocaris. Белые стрелки указывают на сегментарные мышцы, черные — на железистые выросты кишечника. F — Leanchoilia. Околоротовые придатки длинные и разветвленные, эти ветви служат щупами. G — Perspicaris, обладатель двустворчатого головного щита. H, I, J — Helmetia, Sidneyia и Emeraldella, вероятные родственники трилобитов. Длина масштабной линейки 5 мм на A, 10 мм на B—F и H—J и 3 мм на G. Иллюстрация из обсуждаемой статьи в PNAS
Эволюционный процесс, охвативший «стволовых эуартропод», хорошо описывается введенным Александром Георгиевичем Пономаренко термином «артроподизация» (А. Г. Пономаренко, 2004. Артроподизация и ее экологические последствия). Это означает, что признаки членистоногих возникали параллельно во многих (хотя и родственных) эволюционных ветвях, подчиняясь, с одной стороны, общей наследственности, а с другой — общим требованиям окружающей среды. При этом одни и те же адаптации зачастую реализовались в разных эволюционных линиях, во-первых, в разной последовательности, а во-вторых — слегка различными способами. Именно это и дает возможность отличить постепенную параллельную эволюцию от гипотетического явления нового типа сразу «во всеоружии», — подобно Афине, которая вышла в полном вооружении из головы Зевса.
Например, некоторые исследователи считают, что как минимум в двух разных ветвях «стволовых эуартропод» членистые хватательные околоротовые придатки, функционально сходные между собой, на самом деле принадлежат к разным сегментам головы — а значит, развились независимо и никак не могут быть гомологами друг друга (D. A. Legg, J. Vannier, 2013. The affinities of the cosmopolitan arthropod Isoxys and its implications for the origin of arthropods). Это очень правдоподобно: именно так и должен был бы выглядеть процесс артроподизации, если исходить из теоретических представлений, подтвержденных данными по другим группам организмов, которые претерпели аналогичную эволюцию (классический пример такой группы — зверообразные, предки млекопитающих).
«Кроновые эуартроподы»
Для полноты картины следует добавить, что эуартроподы в классическом «рэй-ланкастеровском» смысле более или менее соответствуют современному понятию «кроновая группа эуартропод», или просто «кроновые эуартроподы» (crown-Euarthropoda). Кроновой группой (crown group) в современной систематике называется часть эволюционного древа, охватывающая ближайшего общего предка всех доживших до наших дней членов данной группы вместе со всеми его потомками (неважно, ныне живущими или вымершими; см. Древнейшие предки кольчатых червей могли быть похожи на брахиопод, «Элементы», 26.02.2016). А стволовой группой (stem group) называется та часть древа, ветви которой до современности не дотягиваются (см. рис. 2).
Есть одна группа членистоногих, которую мы еще ни разу не упомянули. Это трилобитообразные (Trilobitomorpha). Трилобиты — широко известные палеозойские животные, в целом похожие внешне на мокриц, но обладавшие огромным разнообразием как облика, так и образа жизни (см. Р. Форти, 2014. Трилобиты. Свидетели эволюции). У них есть четкое деление тела на тагмы, а также настоящие антенны и двуветвистые членистые конечности. Все это — эуартроподные признаки. Современные биологи склонны относить трилобитов к кроновой группе эуартропод, несмотря на то, что они полностью вымерли. Правда, к кому они ближе внутри кроновой группы — к хелицеровым или к жвалоносным, — пока непонятно. Молекулярные методы, которые могли бы однозначно решить этот вопрос, в данном случае неприменимы: ведь последние трилобиты исчезли в пермском периоде, и никакой ДНК от них не осталось.
Первые членистоногие
Теперь мы узнали вполне достаточно, чтобы оценить по достоинству крайне интересный обзор, опубликованный весной 2018 года коллективом из четырех довольно молодых, но уже известных палеонтологов (один соавтор родом из Канады, трое других — из Британии). Все они — специалисты по ископаемым членистоногим, их родственникам и их происхождению. Главный вопрос, который интересовал авторов: что могут нам сообщить эуартроподы (как «кроновые», так и «стволовые»), если рассматривать их в качестве свидетелей кембрийского взрыва?
Чтобы вникнуть в суть дела, нам понадобится немного цифр. По наиболее свежим датировкам, кембрийский период начался 541 миллион лет назад. Остатки членистоногих появляются в кембрийских отложениях отнюдь не сразу. Первые найденные в ископаемом состоянии эуартроподы «кроновой группы» — это трилобиты, самые ранние остатки которых имеют возраст 521 миллион лет. Причем обнаруживаются они почти одновременно в нескольких точках Земли: едва появившись, трилобиты быстро приобрели глобальное распространение (очевидно, не в последнюю очередь потому, что в жизненном цикле у них были планктонные личинки, которых могут свободно переносить морские течения). Что касается «стволовых эуартропод», то самый древний их представитель — существо, относящееся к группе аномалокаридид, от которого найден характерный околоротовой придаток (A. C. Daley, D. A. Legg, 2015. A morphological and taxonomic appraisal of the oldest anomalocaridid from the Lower Cambrian of Poland). Возраст этой находки пока определен лишь приближенно: 519–520 миллионов лет. Чуть позже, 518 миллионов лет назад, на сцену выходит так называемая Чэнцзянская биота, где «стволовые эуартроподы» представлены во всем великолепии. И наконец, 514 миллионов лет назад в летописи появляются микроскопические остатки первых ракообразных. Еще через несколько миллионов лет ракообразных становится много — фауна приближается к современной, кембрийский взрыв остается позади.
Что дают нам все эти даты? Прежде всего, налицо парадоксальный факт: «стволовые эуартроподы» появляются в палеонтологической летописи позже, чем «кроновые». Грубо говоря, предки появляются в летописи позже потомков (пусть и совсем ненамного позже). Что это значит? Никто, конечно, не сомневается, что «кроновые эуартроподы» произошли от кого-то из «стволовых». Но не менее несомненно и то, что какое-то время эти группы существовали вместе. Скорее всего, дело в том, что «стволовые эуартроподы» в принципе хуже представлены в летописи: их кутикула была существенно менее твердой, чем у трилобитов и ракообразных, а потому их тела (или части тел) с меньшей вероятностью сохранялись в донных осадках. Тут стоит вспомнить, что у онихофор, которых не зря называют «бархатными червями», кутикула до сих пор мягкая, бархатистая, растягивающаяся, — совсем не такая, как у ракообразных или насекомых.
Есть, правда, и другая возможность. Что, если трилобиты — это не «кроновые эуартроподы», а одна из групп «стволовых», вышедшая на «истинно эуартроподный» эволюционный уровень совершенно самостоятельно? Это теоретически не исключено, но нуждается в серьезных доказательствах, которых пока нет.
Как видим, в первые двадцать миллионов лет кембрия ни «стволовые», ни «кроновые эуартроподы» не обнаруживаются (во всяком случае, пока — не стоит забывать, что палеонтология продолжает стремительно развиваться и новые находки могут возникнуть в любой момент). Значит ли это, что они появляются в летописи внезапно? Нет. Как обычно и бывает в таких случаях, находкам захороненных тел (или отпечатков тел) предшествуют находки ископаемых следов, предположительно оставленных теми же организмами. Это тоже достойный предмет изучения. Существует целая наука об ископаемых следах — палеоихнология. Итак, в отложениях начала кембрия присутствуют довольно многочисленные ископаемые следовые дорожки, часть из которых, видимо, принадлежит «стволовым эуартроподам», а часть «кроновым». Возраст древнейших из них — примерно 537 миллионов лет.
Ну, а что было еще раньше? Весь отрезок истории Земли, предшествующий началу кембрийского периода, принято называть докембрием. Непосредственно с кембрием снизу — со стороны более древних времен — граничит эдиакарский период, относящийся к протерозойской эре. Живая природа эдиакария уникальна. Она резко отличается от живой природы кембрия. Следы членистоногих — «стволовых», «кроновых» и вообще каких бы то ни было — в эдиакарии, по словам некоторых авторов, «поразительным образом отсутствуют» (M. G. Mangano, L. A. Buatois, 2014. Decoupling of body-plan diversification and ecological structuring during the Ediacaran—Cambrian transition: evolutionary and geobiological feedbacks). Впрочем, это не так уж и поразительно, если исходить из вполне вероятной гипотезы, что членистоногих тогда просто не было.
Загадочная сприггина
Однако голым отрицанием тут не отделаешься. Существует такая вещь, как тафономия — наука о закономерностях захоронения ископаемых остатков, которой дал название великий Иван Антонович Ефремов. Каждый палеонтолог прекрасно знает, что отсутствие той или иной группы организмов в палеонтологической летописи может объясняться эволюционными причинами (эта группа либо еще не возникла, либо уже вымерла), а может и тафономическими (эта группа существовала, но жила в таких условиях, где захоронение остатков ее представителей было крайне маловероятно). Выбор между этими вариантами часто приводит к серьезным дискуссиям. Здесь перед нами именно такой случай. Что, если отсутствие в летописи докембрийских членистоногих — это на самом-то деле тафономический артефакт? Такое возможно, если условия захоронения в эдиакарии были принципиально иными, чем в кембрии.

Сприггина
Понимая эту возможность, авторы обзора дотошно проанализировали данные о том, насколько эдиакарские местонахождения древних организмов (а их известно не так уж мало) отличаются от кембрийских. Был сделан вывод, что наиболее важные с точки зрения нашей проблемы типы сохранности являются для эдиакария и кембрия фактически общими. Речь идет не только о классических лагерштеттах «типа Бёрджесс», но и о захоронениях фосфатизированных микрофоссилий «типа Эрстен» (Orsten-type), которые тоже дают очень интересные находки. Эти типы местонахождений отнюдь не ограничиваются кембрием: в эдиакарии они тоже распространены, и в них отлично сохраняются тогдашние организмы, причем весьма разнообразные. Иное дело, что сплошь и рядом такие находки бывает сложно интерпретировать: увы, эдиакарские живые существа слишком необычны по современным меркам, и поминавшийся нами принцип актуализма начинает тут давать сбои (см., например: Ископаемые эмбрионы из Доушаньтуо, вероятно, не эмбрионы, «Элементы», 09.01.2012). Но уж если бы там были членистоногие — то они сохранились бы наверняка, по крайней мере, хоть в каком-нибудь количестве.
Более того, можно утверждать, что по совокупности геохимических факторов, влиявших на захоронение ископаемых, кембрий был ближе к эдиакарию, чем к более поздним эпохам. Именно к концу кембрия условия в морях сильно изменились — в первую очередь из-за деятельности животных, которые связывали растворенные соли в материале своих раковин, меняя химический состав морской воды, а заодно активно фильтровали эту воду, делая ее более прозрачной и тем самым улучшая условия для водорослей, насыщающих океан кислородом. Кроме того, животные вовсю рыли норы в морском дне, перемешивая грунт и в результате обогащая кислородом его тоже. Все это, вместе взятое, сокращало возможности для образования стабильных бескислородных минерализованных осадков — а как раз из таких осадков и получаются лагерштетты. Неудивительно, что после кембрия их стало гораздо меньше.
Итак, объяснить отсутствие докембрийских членистоногих чисто тафономическими причинами невозможно. Правда, известно несколько эдиакарских ископаемых, которых разные авторы объявляли членистоногими (или членами вымерших типов, близких к ним). Но эта гипотеза ни в одном случае не подтверждена сколько-нибудь надежно.
Типичным примером такого ископаемого можно назвать сприггину (Spriggina) — эдиакарское существо с вытянутым сегментированным телом и неким подобием головного щита. Ее не раз описывали то как кольчатого червя, то как членистоногое (см., например: M. McMenamin, 2003. Spriggina is a trilobitoid ecdysozoan). Однако, во-первых, у сприггины не найдено никаких усиков, ножек, челюстей и вообще конечностей. А во-вторых, обычную для животных двустороннюю симметрию у нее заменяет симметрия скользящего отражения, когда правая и левая стороны тела сдвинуты относительно друг друга примерно на половину сегмента. Уже одно это означает, что если сприггина и была многоклеточным животным, то от плана строения членистоногих ее план строения отличался радикально. Принадлежать к этому типу она никак не могла. Примерно так же обстоит дело и со всеми остальными эдиакарскими находками.
История со сприггиной хорошо показывает, насколько усложняется работа палеонтологов при погружении в древнейшие времена, где уже не действуют сравнения с современной флорой и фауной. Ее предполагаемая более поздняя родственница — метасприггина (Metaspriggina) — в итоге вообще оказалась хордовым животным (P. Janvier, 2015. Facts and fancies about early fossil chordates and vertebrates). Ну, а сама сприггина пока что так и остается загадкой.
Ошибка великого Лайеля
Итак, сумма данных по древним членистоногим на сегодняшний день дает следующую непротиворечивую картину: общий план строения членистоногих (и шире — панартропод) сложился примерно на рубеже эдиакария и кембрия. Успех этого плана строения привел к адаптивной радиации — довольно быстрому разделению группы на множество самостоятельно эволюционирующих ветвей, в которых начался процесс артроподизации. Примерно через 20 миллионов лет этот процесс завершился появлением «истинно эуартроподной» модели, обладатели которой — они же «кроновые эуартроподы» — с тех пор и господствуют в биоте Земли.
Противостоящая этой точке зрения гипотеза «долгой скрытой докембрийской эволюции» была основана в первую очередь на методе молекулярных часов (см. Диверсификация животных началась задолго до кембрийского взрыва, «Элементы», 13.12.2011). Сравнительный подсчет аминокислотных и нуклеотидных замен раз за разом приводил исследователей к выводу, что расхождение главных эволюционных стволов животных случилось задолго до кембрийского взрыва — в глубоком докембрии. Согласно одной из первых таких работ, отделение членистоногих от других животных должно было произойти 1200 миллионов лет назад (A. G. Wray et al., 1996. Molecular evidence for deep Precambrian divergences among metazoan phyla). Сейчас ясно, что подобные оценки совершенно нереальны. Большое количество нуклеотидных замен, отличающих эволюционные ветви друг от друга, может быть связано не только с древностью расхождения этих ветвей, но и со скоростью их эволюции после расхождения. Чем выше эта скорость, тем больше замен могло накопиться за тот же промежуток времени. Поэтому молекулярные часы обязательно нужно калибровать по палеонтологическим данным, как это и делается в современных работах (см. Членистоногие подтверждают реальность кембрийского взрыва, «Элементы», 17.11.2013). Одна из таких работ, например, датирует расхождение жвалоносных и хелицеровых интервалом 514–536 миллионов лет назад (M. dos Reis et al., 2015. Uncertainty in the timing of origin of animals and the limits of precision in molecular timescales). А это уже прекрасно согласуется с непосредственными свидетельствами ископаемых остатков.
Приведем имеющий косвенное отношение к нашей теме факт из истории науки. Величайший геолог XIX века Чарльз Лайель (Sir Charles Lyell), с именем которого принято связывать многократно здесь упоминавшийся принцип актуализма, объяснял отсутствие остатков млекопитающих в палеозойских породах, как бы мы сейчас сказали, тафономическими причинами: мол, млекопитающие живут на суше, а захоронение костей происходит в первую очередь в морских осадках, вот они и не дошли до нас с тех пор (A. Hallam, 1998. Lyell’s views on organic progression, evolution and extinction). Лайель, безусловно, был великим ученым, но в данном случае он ошибся. Сейчас мы точно знаем, что никаких млекопитающих в палеозое не существовало: они появились на целую геологическую эру позже. Лайелю, научное мировоззрение которого было основано на убеждении в глубоком постоянстве факторов и процессов, действовавших на поверхности Земли в течение всей ее истории, было слишком трудно представить себе возникновение на этой планете чего-то совершенно нового — например, нового класса животных. Можно сказать, что это была своеобразная разновидность экономии мышления.
Дискуссия вокруг гипотезы «долгой скрытой докембрийской эволюции» невольно вызывает в памяти эту ошибку великого Лайеля. Впрочем, тут есть оговорки. Во-первых, возникновение самых крупных эволюционных стволов животных вполне может датироваться докембрием, пусть и неглубоким. Во-вторых — и это самое главное — возникший конфликт между гипотезами стопроцентно разрешим. Для этого нужны всего лишь факты. Палеонтология докембрия сейчас развивается быстро. Даже одной-единственной надежно подтвержденной находки докембрийского членистоногого будет достаточно, чтобы противники гипотезы «долгой скрытой докембрийской эволюции» сразу изменили свои взгляды. Посмотрим, произойдет ли это.
Источник: Allison C. Daley, Jonathan B. Antcliffe, Harriet B. Drage, and Stephen Pates. Early fossil record of Euarthropoda and the Cambrian Explosion // Proceedings of the National Academy of Sciences. 2018. V. 115. № 21. P. 5323–5331. DOI: 10.1073/pnas.1719962115.
Сергей Ястребов, Элементы.ru


