

Изучение большой коллекции ископаемой пыльцы и листьев позволило международному коллективу палеоботаников проследить историю тропических дождевых лесов Колумбии в конце мелового и начале палеогенового периодов.
Исследование показало, что великое вымирание на рубеже мела и палеогена привело к радикальной перестройке лесной экосистемы. Позднемеловой дождевой лес сильно отличался от современного: он не имел сомкнутого полога и сложной многоярусной структуры, в нем было много хвойных деревьев, папоротников и узкоспециализированных насекомых-фитофагов. На рубеже мела и палеогена видовое разнообразие растений резко снизилось и вернулось к докризисному уровню лишь спустя 6 миллионов лет. Новые дождевые леса, выросшие на месте прежних во второй половине палеоценовой эпохи, были по своей структуре и таксономическому составу больше похожи на современные, чем на позднемеловые. Это нельзя объяснить климатическими изменениями, потому что климат в регионе остался прежним. К предполагаемым причинам экосистемной перестройки исследователи относят избирательное вымирание хвойных (араукариевых), гибель крупных фитофагов (растительноядных динозавров) и обогащение почвы фосфором и азотом в результате падения Чиксулубского метеорита и экспансии растений семейства бобовых.
66 миллионов лет назад, на рубеже мела и палеогена, произошло одно из самых катастрофических массовых вымираний в истории земной жизни. Чтобы понять его причины, тысячи геологов и палеонтологов десятилетиями изучали пограничные мел-палеогеновые отложения по всему миру. Эти усилия не пропали даром. На сегодняшний день можно считать доказанным, что главной причиной катастрофы было падение Чиксулубского астероида (см.: Жизнь вернулась в кратер Чиксулуб почти сразу после падения астероида, «Элементы», 08.06.2018; Доказана связь кратера Чиксулуб с глобальной иридиевой аномалией на границе мела и палеогена, «Элементы», 16.03.2021). Свою роль, пусть и второстепенную, сыграла также вспышка вулканической активности на территории современной Индии (Радиоизотопные датировки подтвердили связь между падением Чиксулубского метеорита и усилением траппового вулканизма, «Элементы», 05.10.2015; Основная причина мел-палеогенового вымирания — падение астероида, а не формирование Деканских траппов, «Элементы», 05.03.2020). Массовое вымирание не было закономерным итогом предшествующей эволюции биоты, как предполагали некоторые палеонтологи. Однако эта эволюция во многом предопределила, какие группы переживут кризис, а какие вымрут. Например, птицы вытеснили птерозавров из мелкого размерного класса (см.: Полет птерозавров в ходе эволюции неуклонно совершенствовался, «Элементы», 02.11.2020), а у крупных активных животных было мало шансов пережить суровую голодовку, вызванную коллапсом пастбищных пищевых цепей.
Считается, что пыль, сажа и сульфатные аэрозоли, поднятые в воздух падением астероида, на месяцы или годы затмили солнечный свет. Это привело к резкому снижению первичной продукции фотосинтетиков. В такой ситуации наилучшие шансы на выживание были у всякой мелочи, способной продержаться за счет детритных пищевых цепей, а также у терпеливых тварей вроде крокодилов, спокойно относящихся к многомесячным голодовкам.
Давно и хорошо известно, что катастрофа сильно ударила по наземной и морской фауне и фитопланктону. То, что наземная флора тоже подверглась массовому вымиранию, стало выясняться лишь относительно недавно (D. J. Nichols, K. R. Johnson, 2008. Plants and the K-T Boundary). В отличие от многих групп животных, у растений массовое вымирание заметно лишь на видовом уровне, но не на уровне старших таксонов (например, семейств). Скорее всего, это связано с экологическим разнообразием семейств растений и широким перекрыванием их адаптивных зон. Поэтому глобальные палеонтологические базы данных образца последней четверти XX века, содержавшие обобщенные и довольно-таки грубые данные по целым семействам или в лучшем случае родам, создавали обманчивое впечатление, будто наземные растения (а также насекомые) пересекли рубеж мела и палеогена практически без потерь.
Эта иллюзия была развенчана в ходе скрупулезного изучения ископаемых спор, пыльцы и листьев из пограничных мел-палеогеновых отложений Северной Америки. Самый характерный маркер массовой гибели флоры — резкий пик относительного обилия спор папоротников в отложениях, залегающих непосредственно над пограничным слоем со следами Чиксулубского импакта. В позднемеловых лесах было много папоротников, которые, по-видимому, были важными компонентами ранних (пионерных) стадий сукцессии, то есть первыми разрастались на участках, по каким-то причинам лишившихся растительного покрова. «Папоротниковый пик» сразу над границей мела и палеогена обнаружен во многих регионах помимо Северной Америки, но без детальных североамериканских данных его было трудно правильно интерпретировать.
В последние годы стали появляться подробные данные, показывающие резкую смену растительных сообществ в других регионах, например, в Патагонии и Новой Зеландии. По-видимому, в разных местах растительность пострадала в разной степени. Например, в северной части Великих Равнин вымерло около 30% палиноморф (то есть видов пыльцы: соответствие между разными частями ископаемых растений установить удается редко), а докризисный уровень разнообразия растений и насекомых-фитофагов (о них судят по повреждениям на листьях) восстановился лишь к концу палеоцена, то есть через 10 млн лет после массового вымирания. В Патагонии и Новой Зеландии безвозвратно исчезло лишь 10% палиноморф, и разнообразие восстановилось гораздо быстрее.
Статья большого международного коллектива палеоботаников, палинологов и палеоэнтомологов, опубликованная недавно в журнале Science, подводит итоги многолетнего изучения маастрихтских и палеоценовых флор Колумбии. Статья проливает неожиданно яркий свет на историю дождевых тропических лесов — уникального биома, в котором сконцентрирована большая часть современного биоразнообразия (рис. 1). Происхождение этих лесов — давняя научная загадка (см.: В. В. Жерихин, 1993. История биома дождевых тропических лесов). Исследование показало, что дождевые тропические леса современного типа (по крайней мере неотропические, то есть центрально- и южноамериканские) обязаны своим появлением мел-палеогеновому вымиранию.
Изученный материал происходит из 39 точек (геологических разрезов) и охватывает временной интервал от 72 до 58 млн лет назад, то есть весь маастрихтский век и почти всю палеоценовую эпоху. Изучено 637 проб, в которых нашлось в общей сложности 1048 видов пыльцы и спор. Общее число «нахождений» (occurences), то есть комбинаций «место/геологический возраст/вид пыльцы», — 53 029. Кроме того, изучены ископаемые листья из трех хорошо сохранившихся древних флор: одной позднемаастрихтской (формация Гуадуас) и двух позднепалеоценовых (формации Богота и Серрехон), в общей сложности 6950 листьев. Изученный район в маастрихте и палеоцене располагался недалеко от палеоэкватора, примерно в 1500 км к югу от места падения Чиксулубского метеорита, и представлял собой приморскую низменность. В течение всего рассматриваемого интервала времени климат здесь оставался примерно одинаковым, теплым и влажным.
Нужно пояснить, что в данной работе изучались только континентальные (а не морские) отложения. Они формировались непосредственно в тех местах, где росли соответствующие флоры, поэтому по ним можно судить о структуре конкретных растительных сообществ. Однако континентальная ископаемая летопись в принципе более фрагментарна, чем морская, а пробы труднее датировать. Пыльца из морских отложений имеет то преимущество, что ее можно датировать очень точно (например, по фораминиферам и известковому нанопланктону). Но она неизвестно откуда прилетела, и к тому же не всякая пыльца разносится ветром одинаково далеко. Поэтому в обсуждаемой работе нет речи о таких тонких деталях, как непродолжительный послекризисный «папоротниковый пик» (он, впрочем, в Колумбии уже найден в морских отложениях острова Горгонилла, см.: P. R. Renne et al., 2018. Multi-proxy record of the Chicxulub impact at the Cretaceous-Paleogene boundary from Gorgonilla Island, Colombia). Нет в работе и сравнения комплексов пыльцы, с высокой точностью привязанных к последним тысячелетиям мела и первым тысячелетиям палеогена. Для корректной работы с фрагментарным и не очень точно датированным материалом пробы пришлось объединить в 17 групп (time-bins), каждая из которых включает не менее 1000 «нахождений» и дает что-то вроде усредненной характеристики местной растительности за период от полумиллиона до миллиона лет.
Динамика разнообразия растений реконструировалась при помощи статистических методов, разработанных специально для анализа заведомо фрагментарных и неполных палеонтологических данных, с учетом неизбежных различий в представленности реального древнего разнообразия в имеющихся пробах (некоторые из использованных методов описаны в статьях: J. Alroy, 2015. A more precise speciation and extinction rate estimator; D. Silvestro et al., 2019. Improved estimation of macroevolutionary rates from fossil data using a Bayesian framework; J. Alroy, 2018. Dynamics of origination and extinction in the marine fossil record).
Получилось, что разнообразие экваториальной южноамериканской флоры снизилось в раннем палеоцене по сравнению с маастрихтом примерно вдвое, долго оставалось низким и вернулось к докризисному уровню лишь спустя 6 миллионов лет (рис. 2, B). К концу палеоцена оно превысило маастрихтский уровень и продолжало расти вплоть до раннеэоценового климатического оптимума, когда на Земле стояла редкостная жара — градусов на 14 теплее, чем сейчас (см.: Paleocene—Eocene Thermal Maximum).
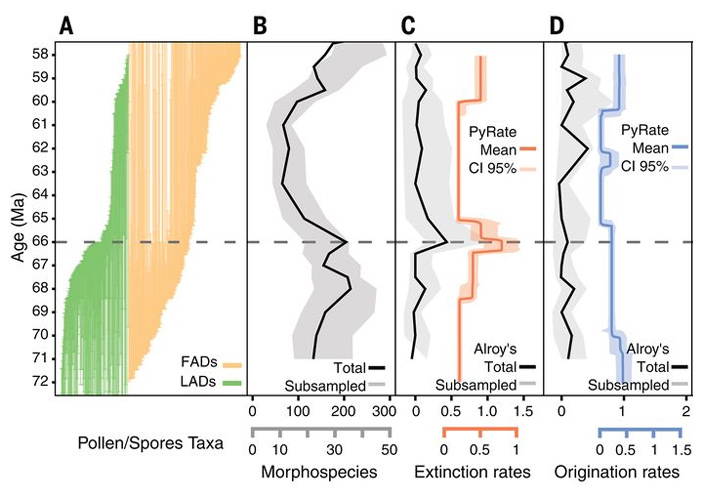
Рис. 2. Динамика разнообразия растений экваториальной Южной Америки в маастрихте — палеоцене по палинологическим данным. По вертикальной оси — время в млн лет назад. Горизонтальная пунктирная линия — рубеж мела и палеогена, когда упал Чиксулубский метеорит. A — интервалы существования споро-пыльцевых «видов» (палиноморф). Каждая тонкая вертикальная линия соответствует одному виду. Зеленым цветом показаны виды, вымершие в рассматриваемом интервале времени, оранжевым — впервые появившиеся в нем. B — реконструированная динамика видового разнообразия. C, D — скорости вымирания и появления видов, оцененные двумя разными методами. Изображение из обсуждаемой статьи в Science
Пик вымирания видов совпадает c границей мела и палеогена (66 млн лет назад) с той точностью, которую могут обеспечить использованные данные и статистические методы. В данном случае это плюс-минус 0,3–0,4 миллиона лет. Фактически в работе утверждается, что пик вымирания приурочен к интервалу от 66,4 до 65,7 млн лет назад. В течение этого интервала средняя скорость вымирания была в 10–15 раз выше фонового уровня, характерного для других интервалов маастрихта и палеоцена. Здесь нужно учитывать, что если вымирание на рубеже мела и палеогена на само деле было мгновенным, то фрагментарность данных и неточность датировок как бы размазали это одномоментное событие на 700 000 лет. В переводе на простой человеческий язык всё это значит, что экваториальная флора Южной Америки подверглась массовому вымиранию на рубеже мела и палеогена — точно так же, как динозавры и множество других групп организмов по всему миру.
Сравнение таксономического состава растительных сообществ при помощи кластерного анализа и других методов показало, что 17 изученных пыльцевых флор (17 групп проб, о которых говорилось выше) четко распадаются на два кластера: маастрихтские и палеоценовые. Флоры конца маастрихта больше похожи на предшествующие маастрихтские флоры, даже отстоящие от них во времени на 5 млн лет, чем на раннепалеоценовые, а те, в свою очередь, больше похожи на позднепалеоценовые, чем на маастрихтские. Это значит, что на рубеже мела и палеогена состав сообществ изменился радикально и необратимо.
В маастрихе пыльца покрытосеменных составляла в среднем 48% от общего числа спор и пыльцевых зерен. В палеоцене эта доля выросла до 84%. Доля спор папоротников уменьшилась втрое, а пыльцы голосеменных стало меньше в шесть раз. Голосеменные (представленные в основном араукариевыми) присутствуют в 75% маастрихтских проб и только в 24% палеоценовых. Таким образом, палеоценовые пыльцевые флоры отличаются от маастрихтских абсолютным преобладанием покрытосеменных и низкой численностью папоротников и араукарий.
Три изученные листовые флоры (позднемаастрихтская и две позднепалеоценовых) включают листья покрытосеменных с небольшой примесью папоротников. Судя по крупному размеру и форме листьев (ровные края, вытянутые кончики для стекания капель), листья принадлежали растениям равнинного тропического дождевого леса (рис. 3).

Рис. 3. Ископаемые листья палеоценового (A—K) и маастрихтского (L—W) тропического дождевого леса. C, E, J — представители семейства бобовых. Изображение из обсуждаемой статьи в Science
Судя по площади листьев (P. Wilf et al., 1998. Using fossil leaves as paleoprecipitation indicators: An Eocene example), количество осадков составляло 234–293 см в год для маастрихтской флоры и 182–184 и 240–308 см в год для двух палеоценовых флор. Это нормальное количество осадков для тропического дождевого леса.
Ранее уже было замечено, что палеоценовые флоры Колумбии похожи по таксономическому составу на современные неотропические дождевые леса. Авторы проверили это при помощи нескольких статистических методов, используя для сравнения различные участки современных дождевых лесов острова Барро Колорадо (Barro Colorado Island, Панама) и национального парка Амакаяку (Amacayacu National Park, Колумбия). Получилось, что обе палеоценовые флоры намного больше похожи на современный неотропический дождевой лес, чем на маастрихтский. В частности, в палеоценовых флорах Колумбии присутствует большинство семейств, доминирующих в современных неотропических дождевых лесах, а в маастрихтской флоре есть лишь немногие из этих семейств (рис. 4).
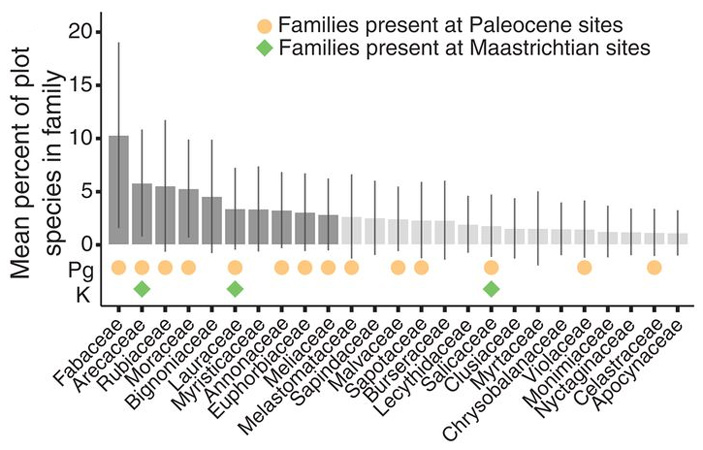
Рис. 4. Семейства растений, к которым относится большинство (75%) видов деревьев в современных дождевых лесах Барро Колорадо и Амакаяку, и их представленность в палеоценовых и маастрихтских флорах Колумбии. Высота столбиков отражает долю видов данного семейства в общем видовом разнообразии. Темно-серым цветом выделены 10 семейств, к которым относится половина всех деревьев (в данном случае не видов, а особей, то есть стволов). Оранжевыми кругами отмечены семейства, присутствующие в палеоценовых флорах Богота и Серрехон, зелеными квадратами — в маастрихстской флоре Гуадуас. Изображение из обсуждаемой статьи в Science
Для современных тропических дождевых лесов характерен сомкнутый полог (рис. 1), острая конкуренция деревьев за свет и сложная ярусная структура, связанная с резким снижением освещенности в нижних ярусах по сравнению с верхними. О наличии сомкнутого полога и ярусности можно судить по внутривидовой изменчивости двух показателей: плотности жилкования (суммарная длина жилок на единицу площади листа; этот показатель зависит от освещенности) и соотношения стабильных изотопов углерода (δ13C; идея в том, что сомкнутый полог затрудняет перемешивание атмосферного CO2 с тем, что выделяется растениями при дыхании). Для маастрихтской флоры характерно унимодальное распределение плотности жилкования и узкий диапазон внутривидовой изменчивости δ13C в кутикуле листьев. Это говорит об отсутствии сомкнутого полога и выраженной ярусности: свет в маастрихтском лесу свободно проходил между кронами деревьев. В палеоценовых флорах наблюдается бимодальное распределение плотности жилкования и большой разброс значений δ13C. Это значит, что палеоценовые дождевые леса Колумбии, как и современные, имели сомкнутый полог и сложную вертикальную структуру. Таким образом, палеоценовые леса были настоящими неотропическими дождевыми лесами современного типа, а маастрихтские сильно от них отличались.
Авторы также изучили повреждения, оставленные на листьях насекомыми-фитофагами. Поврежденные листья попадают в палеонтологическую летопись гораздо чаще, чем сами насекомые. Для таких повреждений разработана особая классификация. На рис. 5 показаны новые типы повреждений, впервые описанные в обсуждаемой статье.

Рис. 5. Повреждения на листьях из маастрихтской флоры Гуадуас (A—F) и палеоценовой флоры Богота (G—H). Изображение из дополнительных материалов к обсуждаемой статье в Science
Разнообразие повреждений оказалось примерно одинаковым в маастрихтской и палеоценовых флорах. Однако нашлось и важное различие: в маастрихтской флоре выше так называемое бета-разнообразие (beta diversity), под которым в данном случае понимается степень несходства видов деревьев по набору повреждений. Это значит, что в маастрихте насекомые-фитофаги были в среднем более специализированными (предпочитали кормиться на ограниченном круге растений), тогда как в палеоцене стало больше фитофагов-генералистов.
Таким образом, позднемеловые дождевые леса Колумбии сильно отличались от палеоценовых и современных. Они не имели плотно сомкнутого полога и сложной ярусной структуры. Наряду с цветковыми в них росли многочисленные папоротники и хвойные (в основном араукариевые) и было много специализированных насекомых-фитофагов.
Леса, выросшие после кризисного рубежа, были похожи на современные по многим признакам: сомкнутый полог, ярусная структура, сходный таксономический состав с резким преобладанием покрытосеменных. Однако общее разнообразие растений в них было меньше, чем в более ранних (маастрихтских) и поздних (эоценовых и современных) лесах. Разнообразие, резко снизившееся на рубеже мела и палеогена, потом очень долго (5–6 миллионов лет) не могло вернуться к докризисному уровню. Ранее похожие тенденции были описаны для палеоценовых флор Северной Америки. Причины столь медленного развития сложных лесных экосистем в начале кайнозоя пока не очень ясны.
Главный вывод работы состоит в том, что дождевые неотропические леса маастрихта и позднего палеоцена сильно отличались друг от друга, хотя климат в регионе не изменился. Светлые маастрихтские леса, лишенные сложной вертикальной ярусности, скорее всего, медленнее фиксировали CO2, меньше испаряли воды и осуществляли менее интенсивный круговорот азота и фосфора по сравнению с лесами современного типа, сформировавшимися в палеоцене.
В связи с этим возникают два вопроса. Во-первых, что мешало маастрихтским деревьям сформировать сомкнутый полог и вертикальную ярусность? Покрытосеменные растения тогда уже были очень разнообразны и, казалось бы, вполне способны создать дождевой лес современного типа. Во-вторых, почему после кризиса не восстановились прежние дождевые леса, а сложились новые?
Авторы предлагают три возможных объяснения. Во-первых, на рубеже мела и палеогена вымерли все крупные фитофаги — растительноядные динозавры, которые в маастрихте еще были весьма разнообразными и многочисленными. Возможно, эти животные, поедая огромное количество растительной биомассы, регулярно прореживали лес, не давали ему сомкнуть кроны и ослабляли конкуренцию между растениями за свет. Может быть, потому и было так много папоротников в маастрихтских лесах, ведь папоротники любят расти на всяких прогалинах и проплешинах.
Вторая гипотеза связана с плодородием почв. Маастрихтский дождевой лес, скорее всего, рос на почвах, бедных азотом и фосфором. Это следует из геологической и климатической истории региона: приморские низменности, существовавшие миллионы лет при неизменном очень влажном климате, должны были растерять биогенные элементы в результате эрозии (в дальнейшем это предположение нужно будет специально проверять). Бедные почвы, возможно, давали некоторое преимущество хвойным, а обилие араукарий не давало лесу сомкнуть полог. Импактные выбросы и пепел от Чиксулубского метеорита, разлетевшиеся повсюду, основательно удобрили землю фосфором. Поэтому на какое-то время главным лимитирующим фактором для растений стал недостаток азота. Это создало благоприятные условия для экспансии бобовых, которые умеют фиксировать атмосферный азот благодаря симбиозу с клубеньковыми бактериями. Действительно, бобовые испытали бурный расцвет сразу после рубежа мела и палеогена, что подтверждается в том числе и молекулярно-филогенетическими реконструкциями. В маастрихтской флоре Гуадуас бобовых нет, а в палеоценовых флорах — есть (рис. 3, C, E, J). Когда почва обогатилась биогенными элементами, быстро растущие покрытосеменные получили преимущество и потеснили араукарии и папоротники.
Третья гипотеза связана с возможным избирательным вымиранием араукариевых на рубеже мела и палеогена. Имеющиеся данные пока не позволяют говорить о такой избирательности как об установленном факте. Однако из общих эволюционно-экологических соображений (изложенных, например, в новости Долгосрочный эволюционный успех обеспечивается не ускоренной диверсификацией, а устойчивостью к изменениям среды, «Элементы», 02.03.2020) вроде бы следует, что экологически однообразные араукариевые должны с большей вероятностью вымирать при резких изменениях среды по сравнению с пластичными покрытосеменными. Так или иначе, после кризисного рубежа хвойные действительно почти исчезли из пыльцевых флор Колумбии. Освободившиеся ниши могли стимулировать дивергенцию покрытосеменных, что в итоге привело к перестройке лесных экосистем.
Для проверки этих гипотез нужны дополнительные исследования. Но в любом случае получается, что мел-палеогеновое вымирание сыграло решающую роль в формировании дождевых тропических лесов современного типа — самого разнообразного биома всех времен.
Источник: Mónica R. Carvalho, Carlos Jaramillo, Felipe de la Parra, Dayenari Caballero-Rodríguez, Fabiany Herrera, Scott Wing, Benjamin L. Turner, Carlos D’Apolito, Millerlandy Romero-Báez, Paula Narváez, Camila Martínez, Mauricio Gutierrez, Conrad Labandeira, German Bayona, Milton Rueda, Manuel Paez-Reyes, Dairon Cárdenas, Álvaro Duque, James L. Crowley, Carlos Santos, Daniele Silvestro. Extinction at the end-Cretaceous and the origin of modern Neotropical rainforests // Science. 2021. DOI: 10.1126/science.abf1969.
Александр Марков, Элементы


