
Известный британский палеонтолог и палеоанималист Марк Уиттон опубликовал в своем блоге первую часть текста, в котором он сравнивает представления о дромеозавридах, созданные масскультурой, с теми, которые составлены на основе современных научных данных. Paleonews.live публикует перевод материала на русский язык.
Реальные рапторы. Поп-культура vs. современная наука
Недавно я поделился в соцсетях фотографией своей борзой Бо, которая стоит рядом со скелетом велоцираптора* . Очень скоро в твиттер посыпались лайки и комментарии.

Многие отмечали, что хотя велоцираптор был карликом в сравнении с собакой, он был грозным хищником, который легко бы пустил Бо на фарш, если бы им когда-либо довелось встретится в реальности. Бедная Бо! Она не заслуживает такой судьбы!
* Вероятнее, что это «велоцирапторин». Если честно, есть некоторая неопределенность в идентификации образца, из которого была отпечатана эта копия.
Комментарии к твиту усилили ощущение, что раптороподобные динозавры мифологизировались в поп-культуре. Во многом это произошло благодаря «Парку Юрского периода», «Краснокожей хищнице» и палеонтологическим публикациям в СМИ 90-х. С тех пор устоялось представление об этих динозаврах, как о быстрых и злобных существах. Обычно в таких случаях вспоминают острые словно бритвы когти, которые были способны разорвать и выпотрошить несчастную жертву; мощные ноги, способные нанести быстрые режущие удары; свирепые укусы и раздирающие плоть зубы, скорость и ловкость гепарда; а также высокий интеллект. От такого описания дейстивтельно остается впечатление, что рапторы были самым ужасными хищниками, которые только когда-либо могли существовать.
Как водится, поп-культура наградила велоцираптора сотоварищи способностями, которые несколько не совпадают с тем, что о них думает наука. Репутация рапторов напоминает то, во что в западной культуре превратились ниндзя. И рапторы, и ниндзя действительно существовали и были способны на завораживающие и устрашающие «подвиги», которые настолько фантастически преувеличены поп-культурой, что это мало напоминает реальность.
Чтобы разобраться на кого эти известные, но «искаженные» поп-культурой, динозавры были похожи, я хочу сравнить расхожие представления о них с последними научными данными. Я не собираюсь снова поднимать тему «Парк Юрского периода неправильно показал велоцирапторов» или обсуждать давно известные вещи, которые вы можете прочесть в другом месте. Например, про несомненное наличие оперения у этих динозавров (Turner et al. 2007; DePalma et al. 2015). Я хочу сосредоточиться на аспектах их образа жизни, биомеханики и экологии. Это обширная тема, поэтому я разделю статью на две части.

В 1993 году голливудский «Парк Юрского периода» познакомил большую часть мира с дромеозавридами. Вероятно, большинство до сих пор представляет, как выглядели и вели себя эти динозавры ориентируясь на фильм. Конечно, множество анатомических особенностей рапторов «Парка» противоречат науке, но сегодня речь не об этом. Фото © Universal, Не знаю точно кто первым залил его в интернет.
Прежде чем начать, давайте быстро обсудим некоторые аспекты терминологии. Во-первых, на какой именно группе динозавров мы сосредоточимся? Велоцираптор входит в состав Dromaeosauridae — группу птицеподобных пернатых теропод. Обычно дромеозавриды характеризуются большими серповидными когтями на вторых пальцах задних лап; длинными негибкими хвостами; узкими и легкими черепами, в челюстях которых находились острые и зазубренные зубы. Окаменелости показывают, что оперение дромеозаврид было в целом сопоставимо с оперением современных птиц. Дромеозавры — это разноплановая группа животных, которая кроме «раптороподобных» форм, также включала в себя узкоголовых и длинноногих уненлангий, мелких и иногда четырехкрылых микрорапторов, а также недавно выделенную группу загадочных гальшкарапторов, которые предположительно вели полуводный образ жизни.
Больше всего рапторам, средних и крупных размеров с большими когтями на ногах, которых мы знаем из массовой культуры, соответствует группа эудромеозавров. Как правило, в эту группу включают несколько родов: велоцираптора, дейнониха, ютараптора, дромеозавра и несколько иных. Клада эудромеозавров признается не всеми палеонтологами, поскольку спорны и не до конца изучены ее точный состав и положение среди остальных дромеозаврид. Тем не менее, это прекрасно соответствует общественному восприятию «рапторов». Более того, это поможет нам в рассуждениях, независимо от того, является ли эта группа истинно таксономической или просто объединяет виды по сходному набору анатомических признаков.
Во-вторых, поговорим о слове «раптор». Примерно два столетия в английском языке это слово было синонимом хищных птиц, но потом все чаще использовалось для обозначения дромеозавров и сходных не-птичьих динозавров. Это произошло после того, как франшиза «Парка Юрского периода» сократила слово «велоцираптор». Некоторые языковые пуристы не любят этот термин (если честно, то я тоже его недолюбливаю), но в палеонтологической литературе все чаще встречается слово «раптор» как разговорное название дромеозавридов. Такие старомодные люди как я, пожалуй, должны признать, что «раптор» «вырос» и охватил всех дромеозаврид. Хотя это усложняет восприятие, когда речь одновременно заходит о хищных птицах и дромеозаврах. Чтобы избежать путаницы, я буду использовать слово «раптор» для обозначения птиц, если не указано иное.
Теперь, когда точки над «i» расставлены, давайте разберемся с тем, насколько представления поп-культуры о эудромеозаврах выдерживают проверку наукой.
Концепция поп-культуры. Эудромеозавры — животные по сути одинаковые, просто разного размера.
Трудно обсуждать дромеозавров в интернете, не обращаясь к реальной таксономической принадлежности велоцираптора, представленного в серии «Парк Юрского периода»*. В качестве фактической основы для созданного на экране существа часто упоминаются дейноних и ютараптор. Такие рассуждения подразумевают, что все эудромеозавры были похожи на дейнониха или велоцираптора. Будто все они были изящными животными с длинными лапами, большими когтями, длинными хвостами и низкими тонкими черепами.
*Уделим немного времени этому столь часто обсуждаемому вопросу. В предисловии романа Роберта Бэккера «Краснокожая хищница» сообщается, что размер велоцирапторов из фильма не был основан ни на чем, кроме желания киношников сделать динозавров зрелищнее. Такое масштабирование дейнониха, а это был самый крупный известный на тот момент дромеозавр, вызывало беспокойство у некоторых участников съемок. «Рапторов» из фильма сделали значительно крупнее, чем велоцираптор и дейноних были в действительности, поскольку реальные размеры выглядели не такими пугающими. Открытие ютараптора немного оправдало кинематографистов, но и тут вышла промашка. Изначально длину ютарапторов оценивали в семь метров, а это существенно превышало размеры динозавров из фильма. Таким образом, «юрские рапторы» размером с человека не имели реального аналога среди дромеозаврид, известных в начале 90-х. На мой взгляд, лучше велоцирапторов из «Парка» рассматривать как усредненных эудромеозавридов, созданных в соответствии с требованиями фильма, без привязки к конкретному роду.

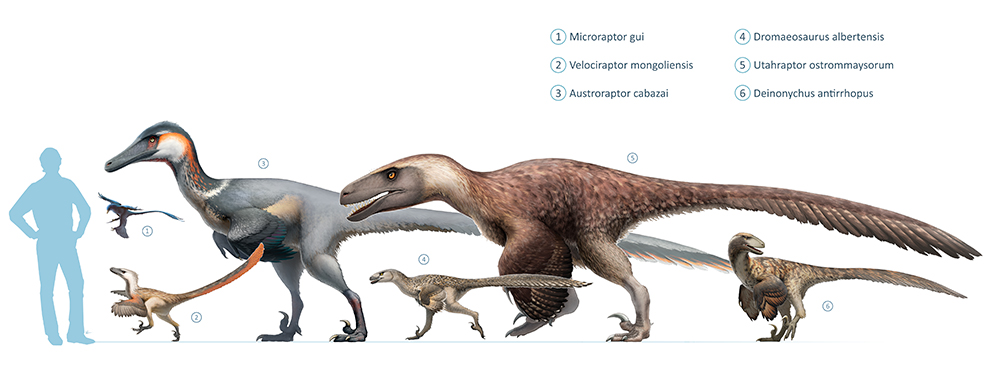
Далеко не все эудромеозавры были «вариациями на тему» велоцираптора. Utahraptor ostrommaysorum находится на другом конце анатомического диапазона. Это крупный хищник, который достигал длины 5-6 метров, а весил 300-500 кг. У него была пропорционально большая голова, мощные лапы и огромные когти. Он напоминал дромеозавра, который стремится вернуться к более традиционному для тероподов строению тела, но не жертвуя некоторым ключевыми адаптациями дромеозаврид.
С начала 90-х научное представление о эудромеозаврах усложнилось. Несмотря на схожее телосложение, они отличались друг от друга длиной лап, пропорциями костей конечностей, размером головы, длиной челюстей, конфигурацией зубов, размерами когтей, гибкостью хвоста и многими другими менее значимыми анатомическими особенностями (e.g. Turner et al. 2012; Paul 2016). Виды эудромеозавров варьировались от мелких животных, едва достигавших полутора метров в длину и (возможно) всего пяти килограммов веса (Bambiraptor) до крупных видов длиной около пяти метров и весом больше 300 кг (Utahraptor). По крайней мере, некоторые из этих крупногабаритных видов, такие как ютараптор и ахиллобатор, были коренастыми существами с крупной головой и сильными челюстями, тяжелыми бедрами, мощными конечностями и большими серповидными когтями. Однако другие крупные эудромеозавры не были такими массивными, а дакотараптор имел схожее строение с дейнонихом и ему подобными, несмотря на то, что был одним из самых внушительных дромеозавров (DePalma et al. 2015). Эудромеозавры меньших размеров тоже отличались разнообразием. Дромеозавр и атроцираптор обладали короткими мордами и крепкими зубами, которые отличались от тонких челюстей дейнониха и ему подобных. Отличался и Adasaurus mongoliensis — это был некрупный эудромеозавр с усиленной задней частью черепа и сильно уменьшенным серповидным когтем (Turner et al. 2012).

Тонкий серповидный коготь адазавра. Иллюстрация: Turner et al. 2012
Эти отличия кажутся незначительными, если учитывать видовое разнообразие теропод в целом. Но они бы стали заметнее и разительнее, если бы мы могли посмотреть на живых динозавров, а не на их окаменелые остатки. Эудромеозавры были не просто велоцирапторами разных размеров, это были виды с различным набором адаптаций и моделей поведения. Эудромеозавры были широко распространенной и долгоживущей группой, и упомянутые различия, вероятно, отражают их способность охотиться на различную добычу, приспособленность к разным экологическим нишам и климату. Вероятнее всего, мы должны рассматривать эудромеозавров как группу, подобную современным группам хищников — кошачьим или хищным птицам, имеющим сходный анатомический и экологический диапазон. В этих группах виды варьируются от небольших охотников на мелких животных до внушительных хищников, которые охотятся на крупную добычу. Как и в случае с кошками или хищными птицами, мы должны быть осторожны, когда обобщаем детали анатомии и внешнего вида дромеозавров.
Концепция поп-культуры . Эудромеозавры были быстрыми как молния и преследовали своих жертв со скоростью, сопоставимой с самыми быстрыми сухопутными животными
Раптор «Парка Юрского периода» неоднократно демонстрировался как невероятно быстрое животное. В фильме он может бежать так же быстро, как и гепард, скорость которого — 109,4–120,7 км/ч. В третьей части фильма мы видели как «рапторы» настигали гадрозавров, догоняли джипы и мотоциклы. Конечно, дейноних был плакатным детищем эпохи «ренессанса динозавров», животным, которое смогло изменить представления о метаболизме, скорости и активности динозавров. Знаменитая реконструкция дейнониха-спринтера Роберта Бэккера — это известный палеоарт, оказавший влияние на многие ученые работы в будущем. Она демонстрирует эудромеозавров как быстрых и проворных существ. Названия классических таксонов эудромеозавров — дромеозавр («бегущая ящерица»), велоцираптор («быстрый вор») — только подчеркивают их стремительность.

Бэккеровский бегущий Deinonychus antirrhopus из работы Острома (1969). Сейчас этой реконструкции уже больше пятидесяти лет , но она до сих пор остается культовой работой эпохи «ренессанса динозавров» и воплощает в себе идею, что динозавры, в том числе дромеозавры, были быстрыми и мощными животными.
Возможно это прозвучит удивительно, учитывая репутацию эудромеозавров, но исследования показывают, что они не были дьявольски быстрыми. Стоит оговорить, что достаточно трудно точно выяснить, как быстро могли передвигаться вымершие животные. Поскольку скорость зависит от ряда факторов, которые нелегко достоверно вычислить по ископаемым находкам. Среди этих факторов масса животного, мышечная масса и скорость, прочность костей, длина шага и многое другое. Дорожки следов могут дать представление о скорости конкретной особи, а такие данные по дромеозаврам известны, но они не могут сообщить нам о максимальной скорости животного. Однако, мы можем оценить скорость животных по пропорциям их конечностей, выстраивая аналогии с современными животными. По таким подсчетам, по крайней мере уже полстолетия как мы знаем, что эудромеозавры не были самыми быстрыми динозаврами (Ostrom 1969; Paul 1988; Kirkland et al. 1993; Carrano 1999; Persons and Currie 2016), несмотря на противоположные утверждения в популярных работах и их трактовку в некоторой научной литературе.
«Замедляет» эудромеозавров то, что в отличие от быстро бегающих животных, у них отсутствуют удлиненные дистальные сегменты конечностей, уменьшена и обтекаемая анатомия пальцев ног, плюсневые кости срослись и сузились. Исследования показали, что задние конечности эудромеозавров, хоть и достаточно мускулистые, но их скорость была снижена ради хватательной анатомии (Ostrom 1969; Fowler et al. 2011).
Изыскания в области биомеханики показывают, что возможности сильно сжимать заднюю лапу и быстро бегать противоположны друг другу. Это что-то вроде развилки на эволюционной дороге, поскольку такие адаптации требуют разной длины конечностей, разного распределения мышц и прочности костей. Близкие родственники эудромеозавров, в том числе уненлагиины и троодонтиды, адаптировались к более быстрому бегу за счет силы ног и, вероятно, были гораздо более проворными и быстрыми существами, чем эквивалентно крупные эудромеозавры (Carrano 1999; Persons and Currie 2016).

Вероятно, эудромеозавры не были самыми быстрыми динозаврами, но они были легкими, мускулистыми и несомненно могли двигаться достаточно быстро в течение короткого промежутка времени, чего было достаточно, чтобы настигнуть добычу.. На рисунке велоцираптор преследует залямблалеста.
Никто всерьез не думает, что эудромеозавры были медлительными существами. Вероятнее всего, они могли двигаться быстро время от времени, просто не могли бежать долго и вряд ли были рекордсменами по скорости. Их жесткие хвосты — четкий признак быстрого передвижения и идеально подходят для стремительной смены направления на большой скорости (Ostrom 1969; Persons and Currie 2012). Логично думать, что эудромеозавры были искусны в засадной охоте, полагаясь на короткий скоростной бросок и ловкость ради поимки добычи. Но вряд ли они были хороши в длительном преследовании.
Некоторые эудромеозавры, подобные крупнотелому ютараптору, вероятно не были особо быстрыми животными. Их задние конечности еще меньше приспособлены к бегу, чем у других эудромеозавров, а хвосты не обладают достаточной жесткостью, что говорит нам об уменьшении ловкости и скорости (Kirkland et al. 1993). Судя по пропорциям, можно предположить, что эти виды «пожертвовали» своей адаптацией к бегу, чтобы облегчить большую массу тела за счёт задних лап и головы. Надеюсь, что по мере того, как мы будем больше узнавать об очень больших эудромеозаврах, мы лучше поймем их локомоцию.
Концепция поп-культуры. У эудромеозавров были невероятно мощные укусы
Идея, популяризированная по крайней мере в романе «Парк Юрского периода», заключается в том, что эудромеозавры, помимо того, что были вооруженными когтями-бритвами суперниндзя, также обладали укусом, который заставил бы аллигатора чувствовать себя неудачником. В этой книге велоцираптор буквально пережевывает стальные прутья в благородной, но в конечном счете тщетной попытке убить одного из самых раздражающих персонажей в истории фантастики, чертова Яна Малкольма. Хотя этот стереотип и не так популярен, как остальные, в ответах на твит «Бо vs. дромеозавр», безусловно фигурировали и комментарии о мощных укусах.

Потрясающе сохранившийся череп Velociraptor mongoliensis демонстрирует низкую, узкую, легко построенную конструкцию, характерную для большинства эудромеозавров. Эти черепа принадлежали не мощным «кусакам», а легким и быстрым животным с зубами, приспособленными к быстрому разрыванию плоти. По Turner et al. (2012).
Я уже писал ранее про силу укуса дромеозавра, поэтому не будем долго останавливаться на этом. Глубокие следы укусов на останках тенонтозавра были приписаны дейнониху и использовались в качестве доказательства его мощного крокодилоподобного укуса рядом авторов (Gignac et al. 2010). Но практически все прочие исследования, в которых рассматривалась прочность черепа и сила укуса эудромеозавров, привели к противоположным выводам (Therrien et al. 2005; Sacamoto 2010;Fowler et al. 2011). Как правило, черепа эудромеозавров представляли из себя легкие конструкции, состоявшие из тонких костей, и поэтому были плохо приспособленны для мощных укусов. По всей вероятности, упомянутые кости тенонтозавра были повреждены укусом другого животного. Therrien et al. (2005) сравнил силу укуса дейнониха с укусом 30-килограммового волка. Это впечатляет только пока мы не вспомним, что дейнонихи весили больше волков в два раза, достигая 80 килограммов. Такое различие, вероятно, обусловлено тем, что псовые адаптированы к разгрызанию костей, в то время как эудромеозавры обладали тонкими зубами и челюстями. Скорее всего они обгладывали кости или заглатывали их целиком, чтобы не повредить зубы. Класть руку в пасть к эудромеозавру вряд ли было бы разумно, но есть множество других животных, которые могли бы укусить вас и посильнее. Неизвестно насколько был силен укус ютараптора и ему подобных. Есть веские основания полагать, что у них были большие и сильные черепа, которые, возможно, допускали большую силу укуса. Но для получения более точных данных необходимо иметь большее количество ископаемых остатков этих гигантских дромеозавров.
Концепция поп-культуры. Эудромеозавры атаковали свою жертву острыми как бритва когтями, глубоко вонзая их в плоть, оставляя длинные кровавые раны
Эудромеозавры, которых я видел в своих детских книжках, как научно-популярных, так и в художественных, обычно изображались с жуткими острыми когтями. Эти когти располагались на лапах таким образом, чтобы убивать добычу наиболее кровожадным и зрелищным способом. Поклонникам палеоарта не нужно напоминать о потоке фильмов и книг 1990-х годов, где динозавры, демонстрировали это: стаи дромеозавров используют свои когти, чтобы карабкаться по телу добычи, разрывая шкуру, чтобы оставить длинные, глубокие раны. Я не сомневаюсь, что именно эти «хищнические» сценарии стали основной причиной того, почему дромеозавры стали любимчиками у поклонников динозавров и общества в целом. Мои собственные детские альбомы были полны кровавых артов, вдохновленных этими идеями.

Страница из комиксов «Эпоха рептилий» 93-го года, изображающая атаку дейнонихов на зауропода. Они нападают, используя свои острые словно бритва когти. Несмотря на то, что это изображение из комиксов, оно довольно близко к представлениям о функциях когтя дромеозавров на то время.
Разрывающие плоть когти дромеозавра имели под собой некоторую фактическую основу. Так общепринято интерпретировали наличие серповидного когтя в середине-конце 20 века (например, Ostrom 1969; Bakker 1986, 1995; Paul 1988; Kirkland et al. 1993). Внушительные, похожие на лезвия когти дромеозавров, большие сгибательные бугорки (часть когтя крепилась к сгибательной мускулатуре) придавали этой концепции достоверность.
Однако сама концепция подверглась критике в последние годы, когда мы начали более подробно изучать образ жизни эудромеозавров, руководствуясь биомеханикой. Достаточно достоверно установлено, что несмотря на узость когтей дромеозавра, они совсем не были похожи на нож, способный разрезать шкуру и мышцы. Коготь в поперечном сечении несколько напоминал вытянутую перевернутую грушу, его внутренняя кромка была узкой, но закругленной (Carpenter 2000; Farlow et al. 2011). Маловероятно, что когти «дополнялись» режущими кератиновыми чехлами. Разве только у них было сочетание рогового чехла и когтей, чего не встречается ни у кого из птиц или рептилий (Carpenter 2000; Manning et al. 2006). Это только основные проблемы для гипотезы «когтей-ножей». Мы хорошо знаем из личного опыта, что разрезать шкуру или мясо без остро наточенного ножа — непростая задача. Предположение, что дромеозавры могли затачивать и поддерживать остроту кромки когтя, выглядит крайне наивно. Кончики когтей можно заточить путем удаления истертых и надломанных краешков кератиной оболочки. Это вам подтвердят любые кошатники, у которых коты портили мебель. Но наточить всю длину когтя без специальной технологии — задача непосильная.

Форма когтя эудромеозавра на иллюстрации Carpenter (2000). Обратите внимание на ширину когтей и отсутствие лезвийных режущих кромок по их внутренним краям.
Еще одна проблема «резательной» гипотезы — это сила, которую эудромеозавры могли передать когтям во время удара ногой. Многие видели художественные работы, изображающие агрессивных дромеозавров, дерущихся ногами. Оказалось, что вытянутые ноги — это самый неудобный вариант для применения таких когтей (Farlow et al. 2011; Bishop 2019). Задние конечности эудромеозавров действительно могли придать силы когтю, но для этого лапы должны были бы быть согнуты (Farlow et al. 2011; Bishop 2019). Общая сила, которую эудромеозавры придавали кончикам когтей, была недостаточно большой в сравнении с силой мышц хищников (Manning et al. 2006; Bishop 2019). Таким образом, даже при оптимальных условиях боя маловероятно, что эудромеозавры обладали достаточной силой, чтобы нанести длинные, глубокие раны другим животным. Ни форма когтей, ни наше понимание механики задних конечностей эудромеозавра не подтверждают то, что он мог использовать когти как «режущее» или «разрывающее» оружие.
Контраргументом может послужить сходство поведения птиц, которые пользуются когтями-шпорами для выяснения отношений внутри вида. Многие виды птиц, в том числе обладатели мощных когтей, наносят друг другу раны во время стычек способом, который не сильно отличается от того, который традиционно приписывался эудромеозаврам. Может, подобные движения, с учетом крупных размеров больших эудромеозавров, не были эффективным средством для убийства добычи? Мне думается, что поведение птиц, которые устраивают подобные поединки, может послужить еще одним доказательством против «теории когтей».
Практически во всех случаях, кроме тех, когда борьба идет совсем уж не на жизнь, а насмерть, и кроме тех, когда ослабленная птица не может сбежать и спастись, такие поединки редко заканчиваются чем-то кроме поверхностных травм. У некоторых видов птиц, в том числе у обладателей больших и кривых когтей (орлы, кариамовые), когти используются не для агрессии, а, например, для коммуникации с потомством. (e.g. Silva et al. 2016). Тот факт, что птицы могут без особого вреда переносить удары когтями, подтверждается также тем, что жестокие люди вооружают бойцовых петухов искусственными металлическими шпорами**. Хоть это и не идеальный аналог, когда речь заходит о эудромеозаврах, «птичья модель» поведения показывает, что наличие больших когтей на мощных лапах не превращает их а жестоких убийц— для разрезания плоти когти малоэффективны.
** Это можно рассматривать как проявление вселенской справедливости. Наращивание боевых шпор бойцовым петухам превращает их в птиц, которые могут быть смертельно опасны для человека. Известно по меньшей мере о трех случаях смерти людей после нападения «усовершенствованных» когтями боевых петухов.

Согнутая левая нога дейнониха (Fowler et al. 2011). Я нахожу это изображение убедительным и в то же время пугающим доказательством мощной хватки, которую дейноних имел при жизни. Обратите внимание на боковой сгиб четвертого пальца, обеспечиваемый шаровидным и гнездообразным суставом на конце плюсневой кости. Масштаб линейки 100 мм.
Итак, если серповидные когти эудромеозавра были предназначены не для того, чтобы резать и рвать, то в чем их смысл? Новый прорывной взгляд на функцию когтей возникает если сосредоточиться на строении лап целиком. Становится понятно, что вся структура стопы адаптирована к прокалыванию и мощному захвату. Когти приспособлены к тому, чтобы прокалывать шкуру (Manning et al. 2009), но плохо приспособлены для разрывания плоти, при этом хорошо впиваются в ткань. (Manning et al. 2006, хотя стоит ознакомиться также с работой Fowler et al. 2011, в которой содержится критика этого исследования). Хорошо сохранившиеся ступни показывают, что эудромеозавры могли сжимать их в «кулак», в котором средние пальцы плотно прижимались к длинными костями стопы, а боковые пальцы смыкались с противоположных сторон (Fowler et al. 2011). По всей видимости, эта хватательная адаптация стала компенсацией за снижение скорости эудромеозавра. Способность собирать заднюю лапу в «кулак» соответствует таковой у многих современных хищных птиц.
Исследователи обращают внимание на многочисленные сходства ног эудромеозавров с ногами современных орлов. Ранее такие аналогии практически не проводились.
Теория, что ноги дромеозавров больше связаны с захватом, чем с «разрезанием», имеет важное значение для нашего представления о экологии этих животных, а также предполагает, что многие из наших традиционных концепций, связанных с нападением на добычу, были маловероятными.
Похоже, что грозные когти этих животных были не совсем такими, как мы когда-то думали, а являлись лишь частью общего механизма захвата жертвы. О том, почему такая стратегия выглядит довольно вероятной — читайте в следующей части этой серии публикаций.
- References
Bakker, R. T. (1986). The dinosaur heresies. William Morrow.
Bakker, R. T. (1996). Raptor red. Bantam.
Bishop, P. J. (2019). Testing the function of dromaeosaurid (Dinosauria, Theropoda) ‘sickle claws’ through musculoskeletal modelling and optimization. PeerJ, 7, e7577.
Carpenter, K. (2000). Evidence of predatory behavior by carnivorous dinosaurs. Gaia, 15, 135-144.
Carrano, M. T. (1999). What, if anything, is a cursor? Categories versus continua for determining locomotor habit in mammals and dinosaurs. Journal of Zoology, 247(1), 29-42.
DePalma, R. A., Burnham, D. A., Martin, L. D., Larson, P. L., & Bakker, R. T. (2015). The first giant raptor (Theropoda: Dromaeosauridae) from the hell creek formation. Paleontological Contributions, 2015(14), 1-16.
Farquhar, C. C. (2017). Commentary: Raptor—Evolution of the Term. Journal of Raptor Research, 51(2), 172-179.
Fowler, D. W., Freedman, E. A., & Scannella, J. B. (2009). Predatory functional morphology in raptors: interdigital variation in talon size is related to prey restraint and immobilisation technique. PloS one, 4(11).
Fowler, D. W., Freedman, E. A., Scannella, J. B., & Kambic, R. E. (2011). The predatory ecology of Deinonychus and the origin of flapping in birds. PLoS One, 6(12).
Gignac, P. M., Makovicky, P. J., Erickson, G. M., & Walsh, R. P. (2010). A description of Deinonychus antirrhopus bite marks and estimates of bite force using tooth indentation simulations. Journal of Vertebrate Paleontology, 30(4), 1169-1177.
Kirkland, J. I., Gaston, R., Burge, D., Kirkland, J. I., & Burge, J. D. (1993). A large dromaeosaur (Theropoda) from the Lower Cretaceous of eastern Utah. Hunteria, 2, 1-16.
Manning, P. L., Payne, D., Pennicott, J., Barrett, P. M., & Ennos, R. A. (2006). Dinosaur killer claws or climbing crampons?. Biology Letters, 2(1), 110-112.
Manning, P. L., Margetts, L., Johnson, M. R., Withers, P. J., Sellers, W. I., Falkingham, P. L., … & Raymont, D. R. (2009). Biomechanics of dromaeosaurid dinosaur claws: application of X‐ray microtomography, nanoindentation, and finite element analysis. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology: Advances in Integrative Anatomy and Evolutionary Biology, 292(9), 1397-1405.
Ostrom, J. H. (1969). Osteology of Deinonychus antirrhopus, an unusual theropod from the lower Cretaceous of Montana. Peabody Museum of Natural History, Yale University Bulletin, 30, l65.
Paul, G. S. (1988). Predatory dinosaurs of the world: a complete illustrated guide. Simon & Schuster.
Paul, G. S. (2016). The Princeton field guide to dinosaurs. Princeton University Press.
Persons IV, W. S., & Currie, P. J. (2012). Dragon tails: convergent caudal morphology in winged archosaurs. Acta Geologica Sinica‐English Edition, 86(6), 1402-1412.
Persons IV, W. S., & Currie, P. J. (2016). An approach to scoring cursorial limb proportions in carnivorous dinosaurs and an attempt to account for allometry. Scientific reports, 6(1), 1-12.
Sakamoto, M. (2010). Jaw biomechanics and the evolution of biting performance in theropod dinosaurs. Proceedings of the Royal Society B: Biological Sciences, 277(1698), 3327-3333.
Silva, A. N., Nunes, R., Estrela, D. C., Malafaia, G., & Castro, A. L. (2016). Behavioral repertoire of the poorly known Red-legged Seriema, Cariama cristata (Cariamiformes: Cariamidae). Rev. Bras. Ornitol, 24, 73-79.
Therrien, F., Henderson, D. M., & Ruff, C. B. (2005). Bite me: biomechanical models of theropod mandibles and implications for feeding behavior. The carnivorous dinosaurs, 179-237.
Turner, A. H., Makovicky, P. J., & Norell, M. A. (2007). Feather quill knobs in the dinosaur Velociraptor. Science, 317(5845), 1721-1721.
Turner, A. H., Makovicky, P. J., & Norell, M. A. (2012). A review of dromaeosaurid systematics and paravian phylogeny. Bulletin of the American museum of natural history, 2012(371), 1-206.


